
VEEN is curated by WSU Viticulture Extension. For questions on articles, or to request to submit an article in future issues, reach out to Michelle Moyer.
This issue of VEEN was originally published as a .PDF in April 2013, and posted online. Some content here will differ from that original issue. The issue is archived, but accessible on the VEEN Archive webpage.
A Note from the Editor
With budbreak teasing us to an early start in 2013, the calm before the “storm” is seemingly short. This period is a great time to catch up on your reading, and prepare for the upcoming growing season. As such, I highly encourage you to pick up a copy of the 2013 Grape Pest Management Guide for Grapes, check out wine.wsu.edu, and of course, enjoy this issue of VEEN.
In this issue, we take a look at the Vineyard Beauty with Benefits project, learn a little bit about how berries balance their water, and get an update on a newly-named disease in vineyards across the USA (and Washington!). We also have an update on the Wine Sensory Lab and a great summary on how the wine matrix can effect tannins.
Sit back, enjoy a barrel sample, and read on!
Michelle M. Moyer
Viticulture Extension Specialist
Assistant Professor of Viticulture
WSU Prosser IAREC
Table of Contents
Connect With Us
V&E Certificate Program
Biological Control Enhanced by Native Plants
By David James, WSU Prosser
For the past two seasons, entomologists at WSU-IAREC, with support from the Washington wine grape industry, Western Sustainable Agriculture, Research and Education (WSARE), and the Northwest Center for Small Fruits Research (NCSFR), have been identifying and counting the beneficial insects attracted native flowering plants growing in and near viticultural areas in eastern Washington.
After examining more than 2500 sticky traps associated with over 100 different plant species, the team has devised a preliminary ranking of these native plants in terms of their ability to attract natural enemies of pests.
The number one ranked plant with the highest average number of all types of beneficial insects was the common sagebrush (Artemesia tridentata; Fig. 1). Flowers of this ubiquitous shrub-steppe plant attracted a mean of 505 beneficial insects per trap; most of these were parasitic wasps, including high numbers of Anagrus wasps, important in the biocontrol of grape leafhoppers.

The number two ranked species was Gray Rabbitbrush (Ericameria nauseousus). The familiar yellow, late summer-autumn flowers of this species attracted an average of 413 beneficial insects per trap with predatory bugs, ladybeetles and bees well represented in addition to parasitic wasps.
Other ‘high-performance’ plants included: Western Clematis (Clematis ligusticifolia), Northern Buckwheat (Eriogonum compositum), Showy Milkweed (Asclepias speciosa, Fig. 2), Yarrow (Achillea millefolium, Fig. 3) and Snow Buckwheat (Eriogonum niveum).


Flowering native plants that attract both a large and a diverse number beneficial of insects may have significant potential as enhancers of natural predator populations and therefore be of benefit to wine grape growers. Wine grape pest management in eastern Washington is largely based on conservation biological control (i.e., using native natural enemies of pests). Thus, cultivation and conservation of native plants that attract predators and parasitoids should improve the efficacy and sustainability of biological control.
The next phase of our project is aimed at identifying and clarifying the benefits of native plants in sustainable pest management for wine grape vineyards. For this, five native plant species that attract a large diversity and number of beneficial insects, and may have value as between-row ground covers in vineyards, have been selected for trial in a WSU wine grape vineyard.
With support from the WSU-BIOAg Program, the selected native plants, including Showy milkweed, Yarrow, Northern Buckwheat, Snow Buckwheat, and Coyote Mint, will be established in the vineyard during spring 2013. Although some supplemental irrigation may be required for establishment, these native plants will not require irrigation in future seasons. Baseline data on pest and beneficial insect populations will be collected during 2013, but evaluation of plant impact on these populations will not be possible until flowering occurs in 2014.
An unusual partner in this research will be the Washington State Penitentiary (WSP) in Walla Walla. As part of an innovative partnership established between WSU and WSP in 2012, selected offenders will be trained in insect identification and used to process leaf and sticky trap samples from the native plant vineyard trial.
The plants chosen for the vineyard trial were selected from a list of 40 to 50 plants shown to have potential as enhancers of biological control in wine grape vineyards. Thus, there are many more plants that need testing in vineyard situations, both as ground covers or refugia plantings. Funding will be sought for this testing both in WSU and commercial vineyards but we encourage vineyard owners to try some of our recommended native plants themselves. A full list of native plants with potential in eastern Washington vineyards for improving pest management will be published later this year.
Aside from the direct benefits to grape pest management, growing native plants in or near your vineyard also benefits the conservation of these plants. Benefits also extend to the other fauna which use these plants as hosts or shelter, such as threatened species of native butterflies and bees.
The majority of plants identified thus far are also important for pollinators and some are also host plants for butterflies. Consequently, in partnership with the Washington Department of Fish and Wildlife, we are establishing a vineyard butterfly project in which we will assist interested growers in fine-tuning their native plant habitat for the dual benefits of better pest management and butterfly conservation.
Clearly, the potential of restoring native habitat to vineyards is considerable both in terms of practical as well as community and aesthetic benefits. The opportunities for capitalizing commercially on these benefits are also self-evident.
Water Economy in Grape Berries
By Yun Zhang, PhD Candidate, WSU Prosser
Water is critical in both viticulture and wine production. Many processes in the plant are dependent on water. As a solvent, water determines the concentrations of all of the compounds (sugars, acids, phenolics, etc.) that are essential in determining fruit quality. At maturity, approximately 70-80% of berry fresh weight in grapes is water [1]. Therefore, one could not overestimate the importance of berry water economy to commercial yield and berry quality.
Four components of berry water economy. Grape berries are storage organs, with growth and ripening that are entirely supported by the water and sugar flows from the vine. Imagining grape berries as a checking account, there are two streams of “direct deposit” in terms of water – xylem and phloem. Xylem is the main pipeline transporting water and minerals into berries. Water flow in the xylem follows the hydrostatic pressure gradient, the same mechanism at work in a garden hose: water always flows “downhill”.
On the other hand, phloem transports sugary water, and it is the only source of sugar to berries. Our grape “checking account” does not just receive deposits, water is also withdrawn as a normal physiological process. Berries are not the only organs on a vine that require water and are always competing with other organs, mainly leaves, for water. When leaves are rapidly transpiring, like during a hot day, the pressure gradient points towards them. Thus xylem flow can be reversed, going from the fruit to the leaves. This is what we call xylem backflow. Water also evaporates (transpires) from the berry surface. Although the rate of berry transpiration is much lower than that of leaves (approximately 100 times less), it is still important, especially late in the season when the water “deposits” from the xylem and phloem into the berry, cease. The four components of the berry water economy are summarized in Figure 1.

Balance in berry water “account” is important. In order to mature normally, grape berries need to balance their water checking account. Unlike our bank checking account, grape berries have a “ceiling”, the skin, which limits the amount of water they can take in. The extensibility of the skin is not limitless, and it declines during ripening [2]. If there is too much water coming in but not enough water going out, the pressure exerted on the skin could crack or split the berries (Fig. 2A). If grape berries are not able to get enough water “deposits” to match the “withdrawals”, shrinkage will occur (Fig. 2B). For example, pre-veraison berries could shrink during hot days, especially with severe water stress that limits water supplies from both the xylem and phloem.
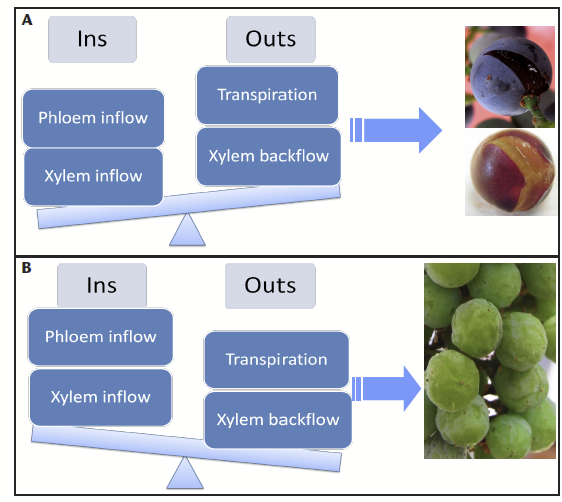
Shrinkage could also happen late in the season with extended hang-time. This kind of shrinkage is due to dehydration, not a physiological disorder (e.g., sour shrivel). The berries shrink because they lose water through xylem backflow and transpiration. Late-season shrinkage (dehydration) may cause substantial yield loss; by the time shrinkage is visible, berries have lost about 10% of their weight. After berries reach approximately 23 °Brix, the phloem stops delivering sugar to berries. Therefore, any increase in °Brix during hang-time is due to dehydration; there is no net gain of sugar content. From one of our experiments, we estimated that Vitis vinifera ‘Merlot’ and ‘Syrah’ berries gained 2.2 and 2.8 °Brix per 10% weight loss, or had 4.7 and 3.9% weight loss to gain 1 °Brix at ripeness. Therefore, it is important to evaluate the cost in yield to reach a certain °Brix level if extended hang-time is required.
Berry transpiration and xylem backflow enhance ripening. There is no doubt that getting stable supplies of water and sugars through the xylem and phloem is essential to the normal ripening of grape berries. However, what roles do berry transpiration and xylem backflow play during berry development? Should we stop berries from losing water to prevent yield loss? In order to answer these questions, we conducted several experiments with three varieties (V. labruscana ‘Concord’, Merlot, and Syrah), to figure out what could happen if we artificially restrict one or both of these pathways.
In a field experiment, we used a commercial anti-transpirant to restrict berry transpiration. Also, to stop xylem backflow, we carefully drilled through the peduncles of clusters to destroy xylem tissue without damaging the phloem. Treatments were applied just before véraison. Both treatments slowed color change and delayed ripening (Fig. 3).

By harvest, berries with either restricted transpiration or xylem backflow had accumulated 33% less sugar compared to untreated ones, and the combination of these two treatments almost doubled the effect (65% less sugar). Besides less sugar accumulation, treated clusters had a much higher cracking incidence than untreated ones (e.g., 7-fold higher with restricted transpiration).
In a separate experiment with potted vines, we used a custom-designed root pressure chamber (Fig. 4) to stop xylem backflow. As before, berry sugar accumulation decreased when xylem backflow was stopped.
The science behind our findings shows that grape berries do not get an unlimited sugary water supply through the phloem. This long-distance transport from leaves (sugar factory) to berries is sophisticated and the “language” of their communication is pressure. Similar to the mechanism in the xylem, yet more complex, phloem sap from the leaves to the berries also follows a pressure gradient.

When phloem delivers sugary water to berries, incompressible water goes into the berries along with unloaded sugars, and pressure is transmitted to the berries. If the pressure cannot be released, as was the case of treated clusters in our experiments, the increased pressure in the berries will become a signal telling the leaves stop sending sugary water. This is why we observed reduced sugar accumulation and delayed ripening in transpiration and xylem backflow restricted clusters. Additionally, if the pressure exerted on the skin exceeds its extensibility, berries will crack, as we saw in our experiment.
It is to an evolutionary advantage that grape berries have more than one pathway to ensure the important sugar delivering process will not be easily disturbed. In our experiments, we have found that berry transpiration varies drastically with changes in environmental conditions: any decrease in air temperature or increase in humidity reduces berry transpiration. Berry transpiration is evidently not a reliable pathway to dispose of excess phloem water, but grape berries have another pathway, xylem backflow, to ensure the continued delivery of sugary water through the phloem and also, to avoid cracking.
To conclude, berry transpiration and xylem backflow are both necessary during berry development. When one or both pathways are restricted, the unbalanced berry water “account” causes delayed ripening, reduced sugar accumulation, and increased cracking.
Irrigation and rainfall. Irrigation and rainfall directly influence berry water economy. Supplying water directly to the soil (e.g., drip irrigation) has different effects than supplying water to the canopy (e.g., overhead sprinkler irrigation or rainfall). When roots take up water from the soil, how much water will be delivered into the berries is controlled by pressure gradients, as described previously.
However, with overhead irrigation or rainfall, water is intercepted by the canopy. This increases canopy and fruit zone humidity, thus decreasing berry transpiration. Water can also move into the berries through their skin [3]. Thus, the berry water economy becomes unbalanced in the overhead irrigation/rain situation, and the risk for berry cracking is increased.
The difference in the effects between supplying water to the soil and to the canopy becomes more dramatic as berries near maturation. As ripening progresses, berries naturally receive less water from the xylem. This means that drip irrigation late in the season does not “pump” water into the berries, thus would not “dilute” berry quality.
Overhead irrigation and rainfall could cause more water to be retained in the berries and even more water to be taken up directly through the skin or stem surface, causing berries to crack. In vineyards with overhead irrigation, late-season irrigation should be approached with caution due to the potential increase in berry cracking.
Avoiding late-season irrigation, however, when it is applied to the soil is unnecessary. Our research results suggest that late-season irrigation does not diluting berry quality when that irrigation is soil-applied. Moreover, soil-applied late-season irrigation should be considered to help replenish soil moisture before the dormant season, thereby reducing cold-damage risk to the vine root system.
References
- Keller M. The science of grapevines: anatomy and physiology. 2010. Burlington, Elsevier Academic Press.
- Matthews M.A., G. Cheng, and S.A. Weinbaum 1987. Changes in water potential and dermal extensibility during grape berry development. J. Am. Society of Horticulture Science 112: 314-319.
- Becker T., and M. Knoche. 2011. Water movement through the surfaces of the grape berry and its stem. American Journal of Enology and Viticulture 62: 340-350.
Update: A New Graft-Transmissible Grape Disease in Washington
By Naidu Rayapati, Sudarsana Poojari, and Olufemi Alabi, WSU Prosser
During the past several growing seasons, we have observed symptoms that look like grapevine leafroll disease (GLRD) in Washington vineyards; however, there were some consistent differences between the symptoms we observed and classic GLRD symptoms (See WSU Extension Bulletin Grapevine Leafroll Disease #EB0762). For example, leaves on these symptomatic vines typically did not show downward rolling of the leaf margins, a characteristic of vines with GLRD. Another difference, seen in red Vitis vinifera cultivars, included small, irregular, red-colored areas between major veins on mature leaves originating from the bottom sections of affected vines which appeared after véraison. The discolored areas on leaves expand with time to become reddish or reddish purple blotches and become strikingly apparent towards end of the season (Fig. 1).

However, there was a wide variation in symptoms across cultivars with red fruit. In some cases, leaves showed red or reddish-purple blotches on leaf margins, and in other cases, the entire leaf showed red or reddish purple discoloration. Even on an affected vine, individual leaves showed differing symptoms. We did not observed any symptoms on V. vinifera cultivars with white fruit or in juice grape varieties (i.e., V. labruscana ‘Concord’).
These symptoms appear to be similar, though not identical, to Grapevine Red Blotch Disease reported in New York and California. It is likely that symptom expression may vary between own-rooted vines planted in Washington vineyards and grafted vines planted in New York and California. Because of this, and the fact that symptoms can vary widely among vines, we are referring to this disease using the umbrella term “Grapevine Redleaf Disease” (GRD), to help ensure that potential symptoms are not overlooked when scouting for this disease.
Understanding Virus Names
When scientific names are used, they are typically presented in italics, such as Vitis vinifera (European wine grape). Italics are used to identify: (i) that the name is in binomial nomenclature (Genus and species), and (ii) that, indirectly, these are the names recognized across the scientific community.
This naming convention, however, is not the same with viruses and viroids. In the virus/viroid world, once the International Committee on Taxonomy of Viruses accepts a name, it is italicized to demonstrate official recognition. For example, Grapevine fanleaf virus is an officially recognized name (hence, in italics). When a virus name is not italicized, it indicates that an official name has not been given to the virus and multiple names may exist. Once an official name is agreed upon, that official name in italics will be used.
What Causes GRD?
One of the first steps in determining whether symptoms are caused by a pathogen rather than an abiotic stress is to see if those symptoms can be transferred from affected to healthy plants. One way to do this is to conduct grafting experiments, where affected (i.e., symptomatic) tissue is grafted to healthy (i.e., asymptomatic) tissue. When budwood derived from symptomatic Merlot and Cabernet franc vines was grafted onto healthy Cabernet franc vines, we saw a transmission of symptoms. This indicated the presence of graft-transmissible pathogens associated with GRD symptoms.
Since GRD symptoms were somewhat similar to GLRD symptoms, we then proceeded to test samples with GRD symptoms for the presence of currently-known Grapevine leafroll-associated viruses (GLRaVs) using molecular diagnostic assays. These test results came back negative; we began to suspect that we were working with either (i) a potential ‘new’ strain of GLRaV, (ii) ‘new’ virus(es) in general, and/or (iii) new virus-like agent(s). Since identifying new disease agents can be difficult using traditional methods, we deployed Next Generation Sequencing (NGS) technology to help in pathogen identification.
The results obtained from NGS revealed presence of a single-stranded DNA virus, tentatively named as Grapevine Redleaf-associated virus (GRLaV) and Grapevine fanleaf virus in grapevines showing GRD symptoms. In contrast, Grapevine rupestris stem pitting-associated virus, Hop stunt viroid, Grapevine yellow speckle viroid 1, Citrus exocortis viroid (CEVd) and Citrus exocortis Yucatan viroid (CEYVd) were present in both symptomatic and asymptomatic grapevines. Molecular and phylogenetic analysis indicated that GRLaV is a new geminivirus with a genome of 3,208 nucleotides (the individual components of a DNA strand) and distinct characteristics from other members of the virus family Geminiviridae. GRLaV is almost identical to recently reported, tentatively named, Grapevine Cabernet franc-associated virus (New York), and Grapevine red blotch-associated virus (California).
How is GRD spread? We have conducted preliminary, greenhouse-based feeding assays to determine how the GLRaV is spread. The results suggest that GLRaV may be transmitted by the Virginia Creeper Leafhopper (Erythroneura ziczac). As a note of caution, these were greenhouse-based studies, and the precise role of a potential vector relationship has yet to be confirmed in a field setting.
How Can GRD be Detected in Plant Tissue?
We have developed a PCR-based method for the detection of GRLaV in both hardwood canes during dormant season and green tissue throughout the growing season. Currently, the role of the different viruses and viroids mentioned above in the production of GRD symptoms is not clear. Since GRLaV was consistently found in vines showing GRD symptoms, this virus has been largely implicated in the production of the associated symptoms. Further studies are in progress to understand the precise role of GRLaV in the development of GRD symptoms.
What are the Potential Impacts of GRD?
To determine whether GRD was negatively impacting vine development, we compared yield and fruit quality from symptomatic to adjacent asymptomatic vines of V. vinifera ‘Merlot’ and ‘Cabernet franc’ in a commercial vineyard. In 2012, GRD-affected Merlot and Cabernet franc grapevines had 22% and 37% less yield compared with corresponding asymptomatic vines. An analysis of fruit quality attributes indicated that berries from GRD-affected vines of both cultivars had lower amounts of total soluble solids (12% less in Merlot, 14% less in Cabernet franc), lower berry skin anthocyanins (4% less in Merlot, 9% less in Cabernet franc), and higher titratable acidity (9% more in Merlot, 16% more in Cabernet franc), compared to berries from asymptomatic vines. In contrast, no significant difference was observed in juice pH. These results indicated a potential negative impact of GRD on yield and key quality attributes.
Conclusion
In summary, using NGS technology, we have identified potential viruses and viroids associated with GRD symptoms. We are also determining the differences between GRD and GLRD in vineyards, in an effort to deploy knowledge-based strategies for the management of both diseases. Finally, based on the success of using NGS technology to identify new viruses and viroids, we are currently developing molecular diagnostic assays for sensitive and specific detection of viruses and viroids in vineyards and for clean plant programs. We hope the development of new information will help prevent the spread of new or newly-identified pathogens, and aid in increasing the awareness of newly-named diseases, such as GRD, among growers and nurseries.
This work was supported, in part, by WSU-ARC, Washington State Grape and Wine Research Program, USDA-Northwest Center for Small Fruits Research, and USDA-NIFA-SCRI (#2009-51181-06027), WSDA-Nursery Assessment Funds, Washington State Commission on Pesticide Registration, and USDA-APHIS National Clean Plant Network. We thank Dr. James Harbertson for advice and Ms. Maria Mireles for technical assistance in biochemical analysis of berry samples, and members of Eureka Genomics for collaboration on the next-generation sequencing project.
WSU Wine Sensory Purchases Electronic Tongue
By Carolyn Ross, WSU Pullman
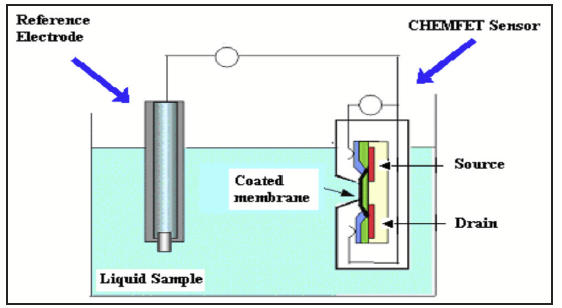
Recently, the Ross Lab purchased a novel piece of analytical equipment, an Electronic Tongue (e-Tongue) by Alpha-MOS (Fig. 1). The e-Tongue is a coated sensor array instrument with an integrated pattern recognition system. It draws on the underlying principle of the neurophysiology of the sense of taste; in other words, how our nervous system translates what we consume into a taste sensation.

The e-Tongue quantifies non-volatile organic and inorganic compounds, and similar to the human tongue, can provide a “taste fingerprint” based on the non-volatile profile of the wine (sweet, sour, salty, bitter, umami, metallic and savory). Using the e-Tongue, we will be able to overlay sensory data and see the influence of that sensory data on wine differentiation.
In a typical operation of the e-Tongue, the sensors interact with the solution via the electrical properties its various chemical components. Our particular e-Tongue uses a potentiometric method of detection which measures a voltage difference between a reference electrode and the various sensors (Fig. 2).
Since the use of the e-Tongue is new, there is not much known how to use it to analyzed wine samples. Our lab has been working on developing these methods to help standardize the analysis process. Using wines that were previously prepared, we separated the different treatments represented in this wines based on their “taste fingerprint” as determined by the e-Tongue. This “taste fingerprint” is assessed based on a discrimination index (DI). The DI is an indication of the ability of the e-Tongue to separate wine samples on their differences in sweetness, bitterness, metallic, spiciness /savory, and sourness responses. A high DI (>80) was observed among the samples.
The responses from the e-Tongue for each wine were then compared to the sensory profiles generated using a trained sensory panel. This comparison showed a strong positive relationship (r >0.93) between the e-Tongue and the sensory panel for the attributes of sweetness, ethanol burn, astringency and bitterness. This indicates that for these attributes the e-Tongue does a good approximation of what a sensory panel would do.
With the rising popularity in wine comes the rising need for sensory research. Unfortunately, using human-based evaluation panels can be costly, time-consuming (training), and ridden with the inherent limitation of subjectivity. The e-Tongue may help alleviate these issues, as it showed great promise in not only discriminating wine treatments, but also as a tool for predicting the sensory attributes of wines. The Sensory Lab at WSU is one of the few research labs currently using this tool for wine analyses. In the future, we plan on using the e-Tongue, in addition to trained sensory evaluation panels, to describe differences between wines.
Thresholds and Rejection of TDN by Consumers
By Carolyn Ross, WSU Pullman
While at Lincoln University in Christchurch, New Zealand for my research sabbatical, my collaborator Roland Harrison and I explored an interesting research question regarding 1,1,6-trimethyl-1,2-dihydronaphthalene (TDN). This compound is important to the aroma profile of a mature Vitis vinifera ‘Riesling’ wine and contributes a ‘petrol’ or ‘kerosene’ note.
In wine, the TDN sensory thresholds, or the lowest concentrations at which the TDN can be detected by humans, have been defined. However, the TDN threshold that affects consumer acceptability of a wine is not clear; these thresholds are typically higher than sensory thresholds. Unfortunately, threshold determinations are notoriously difficult, varying greatly with the methodology used to measure them.
To address this question of where consumer rejection thresholds are for TDN, we developed a protocol that: (i) Determined the threshold of TDN in a given wine, and then, (ii) Used paired preference testing to determine if the consumer would then reject that wine. Both studies were conducted at two locations, NZ (Lincoln) and at USA (WSU-Pullman). Using these two different consumer populations allowed us to determine differences in rejection thresholds of TDN in Riesling wines across different regions/populations. The consumers used in the study (n=36) were self-defined as “regular white wine consumers.”
To test for detection thresholds, 3 samples were given to each panelist, one with a known quantity of TDN, two with no TDN. Panelists were asked to identify which sample was different from the others. This test started with a low concentration of TDN, and then increased that concentration with each presented set of wines. To test for consumer rejection / preference thresholds, each panelist received multiple pairs of Riesling wine samples: one sample was spiked with a known quantity of TDN, one did not contain any TDN. In each of these pairings, participants were asked to indicate which sample was preferred (they did not know which sample was spiked).
The TDN detection threshold value in Riesling for the NZ and USA consumers was 37.9 and 43.2 μg TDN/L, respectively. No significant differences in threshold values were observed between locations or replicate panelist. However, consistent regional differences in TDN preference were found. The consumer rejection threshold for USA was 64 μg TDN/L; the rejection threshold for NZ was 204.8 μg TDN/L. This suggests that consumers in both regions detected TDN at similar concentrations, but prefered / tolerated different levels of TDN. Consumers in NZ tolerated a higher concentration of TDN compared to those in the USA.
These results highlighted the influence of TDN on the acceptance of Riesling wines, and may be useful for those winemakers and marketers wishing to address differences in consumer preferences across regions.
Wine Matrix: Tannin Extraction and Astringency
By Federico Casassa, PhD Candidate, Richard Larsen, and James Harbertson, WSU Prosser
In red wine production, winemakers spend a considerable amount of time both in the vineyard tasting grapes, and in the lab interpreting berry basic analysis (Brix, pH, acidity) and phenolic chemistry (anthocyanins, skin tannins and seed tannins) as harvest approaches. The goal is to assess the optimal fruit “ripeness” for a given wine style.
Tannins play a critical role in this optimal “ripeness”, as they contribute to the oral sensations of astringency, bitterness, and possibly other textural attributes such as wine body and mouthfeel. For many of the solutes presented in the berries (e.g., sugars, acids), a measurement of the amount in the berry provides a fair estimate of the amount that will be extracted during maceration. Tannins, however, seem to be an exception to that rule [1].
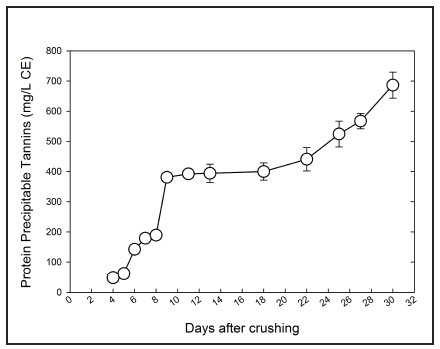
While Figure 1 implies a simple extraction curve of tannins during maceration, true tannin extraction and retention appears to be more complex than implied. Furthermore, the astringency and bitterness sensations winemakers perceive when tasting isolated skins or seeds in the vineyard are perceptually different, both in intensity and duration, from those perceived during wine tasting. These simple observations suggest that major structural changes involving tannins take place during crushing, maceration and wine aging.
Effect on tannin extraction and evolution during winemaking. The extraction of tannins into wine and their fate during post-maceration and aging is governed by the wine matrix composition. Example components of the wine matrix composition include anthocyanins and other compounds such as mannoproteins originating from yeasts, and polysaccharides located in cell walls of mesocarp (i.e., pulp) and berry skin; these components are known to react with wine tannins.
Presence of anthocyanins and formation of polymeric pigments. The presence of anthocyanins during maceration increases the solubility and retention of tannins [2]. This occurs via the formation of polymeric pigments. Polymeric pigments are extremely heterogeneous winemaking artifacts composed primarily of anthocyanins and tannins, but recent research suggests that other wine constituents, such as polysaccharides, mannoproteins, organic acids and carbohydrates, may also be involved in the chemical structure of such pigments [3]. In an experiment in which white wines were produced with different portions of added tannins and anthocyanins, both tannin retention and formation of polymeric pigments increased in the presence of added anthocyanins [2]. This study also found that the stoichiometric addition of anthocyanins relative to tannins approached an ideal proportion; a concentration excess of anthocyanins did not lead to a corresponding increase in pigmented polymer formation. The levels of tannins in both wines were not different, suggesting that while anthocyanin addition increased polymeric pigment formation, it did not increase tannin extraction.
In a separate study, where white wines were produced in contact with skins and seeds also had additions of anthocyanins [4], polymeric pigments and astringency levels were higher in the resulting wines, relative to those observed for the white wine produced with the white pomace alone. Together, these findings suggest that the presence of anthocyanins invariably leads to polymeric pigment formation. However, the proportion of anthocyanins and tannins during maceration will determine the amount of wine tannins that are retained. These hypotheses, however, remain to be tested.
There are scientific reasons, based on both chemical and sensory data, to believe that polymeric pigments confer desirable sensory traits to the final wine. For example, relative to the intact anthocyanins, polymeric pigments are partially resistant to bisulfite bleaching and more resilient to pH changes [5]. This means polymeric pigments confer to the wine a more stable and thus potentially long-lasting color. Beyond the influence of polymeric pigments in color stability, direct sensory evidence of the role of these compounds in the changes in mouthfeel and astringency observed during wine aging is relatively recent. The observed ‘lessening’ of astringency during wine aging was classically thought to be the result of the formation of polymeric pigments involving anthocyanins and tannins of various sizes [6]. However, the structural complexity and heterogeneity of these pigments prevented their isolation and their chemical and sensory characterization. With the advent of new analytical, semi-preparative and preparative high-performance liquid chromatography (HPLC) approaches coupled with nuclear magnetic resonance (NMR), both the chemical and sensory characterization of these compounds has become possible. For example, one study found that polymeric pigments bearing an anthocyanin moiety were less astringent than apple tannins of the same size deprived of anthocyanins [7]. The comparatively lower astringency of polymeric pigments relative to that of intact tannins has been hypothesized to result from an increase in the hydrophilic (water-loving) character of the pigment due to the incorporation of the anthocyanins which have a sugar molecule attached to their structure [2]. This would reportedly decrease the interaction of the tannin pigments with the proteins founds in human saliva, thus reducing astringency. As polymeric pigments were found to contain polysaccharides within their structure, the formation of soluble complexes that do not precipitate with salivary proteins (and hence do not elicit astringency) is also a possibility [8].
Therefore, the empirical observation of decreased astringency in wine aging may not be related to a change in the total amount of tannin present. Rather, the structural modification of wine tannins, primarily resulting from the incorporation of anthocyanins, and, secondarily from the addition of other metabolites such as carbohydrates, proteins, and polysaccharides, may drive the observed changes in perceived astringency during wine aging.
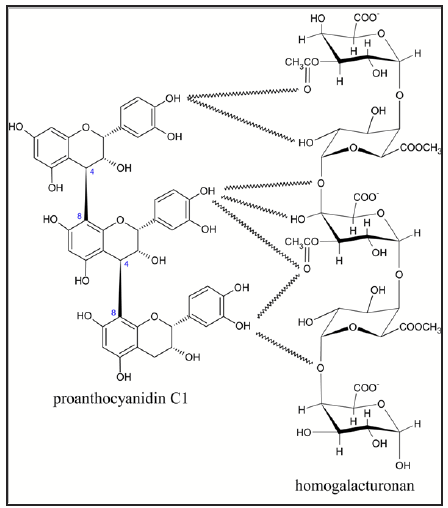
Interaction between proanthocyanidins and cell wall components during maceration. In apple cider production, there is a lack of recovery of high molecular weight tannins that were originally present at the beginning of processing [9]. Chemical interactions between tannins and cell wall material are responsible for this ‘natural fining’ occurring spontaneously during production. These interactions increase with the increasing tannins size [10], and are depicted in Figure 2.
In wines, the failure to recover high molecular weight tannins is thought to be the result of tannin–cell wall interactions that occur during winemaking [11]. Furthermore, a series of studies conducted by the Australian Wine Research Institute showed a significant relationship between the tannin molecular mass and the proportion of tannin adsorbed by skin and pulp cell wall polysaccharides. The end result would be that higher molecular weight tannins are not extractable and/or removed from the wine by interaction with cell wall components during maceration [12,13,14]. If that hypothesis proves true, it would explain why, even though skin tannins are fairly large (between 20 to 30 and up to 80 flavan-3-ols units), the average molecular weight of wine tannins only varies between 2 and 7 units [15,16]. Moreover, these interactions may also explain the post-fermentation extraction of wine tannins during extended maceration. As ethanol can disrupt these interactions, the post-fermentation extraction of tannins observed in Figure 1 after day 20 could be the result of a desorption mechanism mediated by ethanol, which leads to the disruption of these interactions between the extracted tannins that were bound to cell wall material [1, 11]. Studies examining the effect of varying ethanol levels on tannin extraction and evolution under actual winemaking conditions have yet to be carried out.
Extended maceration, polymeric pigments, and missing tannins. Extended maceration increases the amount of tannins retained into wine, and these tannins are mainly derived from the seeds [16,17]. Extended maceration can also promote the formation of polymeric pigments. Experiments conducted in the Research Winery at WSU-IAREC from 2010 to 2012 showed that this enhanced polymeric pigment formation during extended maceration was generally due to the formation of large polymeric pigments (assumed to be composed of more than 3 flavan-3-ol subunits in addition to one or more anthocyanin molecules). However, enhanced formation of small polymeric pigments (assumed to be composed by anthocyanin-tannin adducts of 3 units or less) was also observed.
Interestingly, upon analysis of tannins in fruit, wine and those recovered in the pomace after maceration, it was found that the proportion of tannins that are unaccounted for (i.e., tannins that were present in the fruit but cannot be recovered in the wines or in the skin and seed pomace samples after maceration) was always consistently higher upon application of extended maceration (Fig. 3). In other words, the higher the amount of extracted tannins during extended maceration, the higher the proportion of missing tannins. We have also found that the proportion of unaccounted tannins varies between seasons, vineyards and varieties. While some of these unaccounted tannins may be incorporated into small polymeric pigments during extended maceration, as we had observed, the amount of tannin incorporated into these pigments can only be partially responsible for this comparatively higher proportion of unaccounted tannins [16].
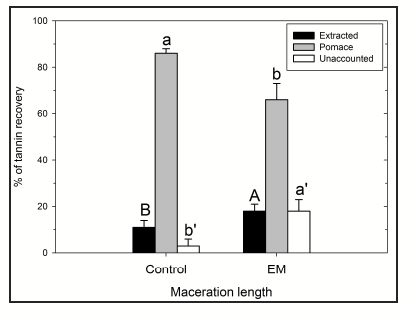
These data suggest that the longer the maceration time, the more pronounced the matrix effect is on sequestering grape tannins. While an additional pool of tannins is indeed extracted during extended maceration, a portion may be irreversibly bound to cell wall components from pulp cells, and to polysaccharides or mannoproteins of grape and/or yeast origin, which are removed after wine settling. The prolonged maceration time may only act by exacerbating this matrix effect.
Conclusions. Tannin extraction into wine is more complex than previously thought. It appears that tannin extraction is governed by a complex relationship between tannin content, anthocyanin extraction (or loss) and polymeric pigment formation during maceration. This points to a more prominent role of the wine matrix, not only as a determinant of the tannin extraction, but also in some sensory properties such as astringency. There is a need to understand the basic chemistry underlying the interaction between tannins, anthocyanins, polysaccharides, and mannoproteins during maceration, as well as understanding the basic chemical effect of fermentation and/or oxidation products such as ethanol and acetaldehyde on wine sensations. An understanding of these reactions will allow scientists to model the evolution of complex wine sensations, such as astringency and bitterness, and ultimately help winemakers to produce wines according to specific stylistic protocols.
Definitions
- Tannins are complex molecules found in the skins, seeds and stems of the grape berries and are composed of individual units named flavan-3-ols. Tannins vary in size according to the number of these units and are typically at higher concentrations in skins (from 10 up to 80 units) than in seeds (from 2 up to 15 units) and wines (from 2 up to 7 units).
- Bisulfite (SO3H–) is a common preservative derived from added sulfur dioxide during winemaking that is able to bleach the wine color through reaction with anthocyanins.
References
- Adams D.O., and R.C. Scholz. 2008. Tannins – the problem of extraction. In Proc. 13th Aus. Wine Ind. Tech. Conf. Blair,pp. 160–164. Aus. Soc. for Vitic. and Oenol., Adelaide, Australia.
- Singleton V.L., and E.K. Trousdale. 1992. Anthocyanin-tannin interactions explaining differences in polymeric phenols between white and red wines. Am. J. Enol. Vitic. 43:63–70.
- Wollman N., and T. Hofmann. 2013. Compositional and sensory characterization of red wine polymers. J. Agric. Food Chem. doi: 10.1021/jf3052576
- Oberholster,A., I.L. Francis, P.G. Iland, and E.J. Waters. 2009. Mouthfeel of white wines made with and without pomace contact and added anthocyanins. Aust. J. Grape Wine Res. 15:59–69.
- Somers T.C., and M.E. Evans. 1977. Spectral evaluation of young red wines: Anthocyanin equilibria, total phenols, free and molecular SO2 and chemical age. J. Sci. Food Agric. 28:279–287.
- Somers T.C.1971. The polymeric nature of wine pigments. Phytochem. 10:2175–2186.
- Vidal S., L. Francis, A.C. Noble, M. Kwiatkowski, V. Cheynier, and E.J. Waters. 2004. Taste and mouth-feel properties of different types of tannin-like polyphenolic compounds and anthocyanins in wine. Anal. Chim. Acta 513:57−65.
- Scollary, G.R., G. Pásti, M. Kállay, J. Blackman, and A.C. Clark. 2012. Astringency response of red wines: potential role of molecular assembly. Trends Food Sci. Tech. 27:25–36.
- Haslam, E. 1998. Practical Polyphenolics: From Structure to Molecular Recognition and Physiological Action. Cambridge University Press, Cambridge, United Kingdom.
- Le Bourvellec C., Guyot, S. and C.M. Renard. 2004. Noncovalent interaction between procyanidins and apple cell wall material. Part I. Effect of some environmental parameters. Biochim. Biophys. Acta 1672:192–202.
- Hanlin, R.L., M. Hrmova, J.F. Harbertson, and M.O. Downey. 2010. Review: condensed tannin and grape cell wall interactions and their impact on tannin extractability into wine. Aust. J. Grape Wine Res. 16:173−188.
- Bindon K.A., P.A. Smith, and J.A. Kennedy. 2010 a. Interaction between grape derived proanthocyanidins and cell wall material. 1. Effect on proanthocyanidin composition and molecular mass. J. Agric. Food Chem. 58:2520–2528.
- Bindon K.A., P.A. Smith, H. Holt, and J.A. Kennedy. 2010 b. Interaction between grape-derived proanthocyanidins and cell wall material. 2. Implications for vinification. J. Agric. Food Chem. 58:10736–10746.
- Bindon K.A., A. Bacic, and J.A. Kennedy. 2012. Tissue-specific and developmental modification of grape cell walls influences the adsorption of proanthocyanidins. J. Agric. Food Chem. 60:9249–9260.
- Hanlin R.L., M.A. Kelm, K.L. Wilkinson, and M.O. Downey. 2011. Detailed characterization of proanthocyanidins in skin, seeds and wine of Shiraz and Cabernet Sauvignon wine grapes (Vitis vinifera). J. Agric. Food Chem. 59:13265–13276.
- Casassa L.F., C.W. Beaver, M.S. Mireles, and J.F. Harbertson. 2013. Effect of extended maceration and ethanol concentration on the extraction and evolution of phenolics, color components and sensory attributes of Merlot wines. Aust. J. Grape Wine Res. 19:25–39.
- Harbertson J.F., M. Mireles, E. Harwood, K.M. Weller, and C.F. Ross. 2009. Chemical and sensory effects of saignée, water addition and extended maceration on high Brix must. Am. J. Enol. Vitic. 60:450–460.